Race is a controversial subject, so I may as well state my views as clearly as possible at the start: 1) there are no significant genetic differences between groups of people around the world, and 2) there are good reasons to believe that we will never find such differences.
It will take some time to explain how I reached these conclusions, but I would like to begin with this perspective. If you have ever wondered about race and genetics, I encourage you to keep reading. You may not agree with my arguments, but I hope you find the discussion informative.
References for all materials are included in the link at the top of the page. Many of the figures are my own. Occasionally, I modified figures from published studies to make them more accessible to readers. In these cases, I indicated that changes were made in the description accompanying each figure.
Why race and genetics?
I decided to create this website for several reasons. I am a science teacher, and my students frequently ask me about race. They ask me about skin color and eye shape. They ask me about sports performance and intelligence. They ask me about human evolution and history. In general, they wonder about differences between people around the world, what these differences mean or do not mean, how they were created, and how they might change in the future.
These subjects are mediated by enduring racist beliefs, grounded in a broad set of false assumptions and conclusions. These beliefs are transmitted, often without question, from person to person, generation to generation.
Part of the reason that racist beliefs persist is that that many people are afraid to speak openly about race. They are hesitant to engage in conversation with others, and they are concerned about how their own views will be interpreted. This is true in social settings, and in the academic world.
For example, despite twenty-five years of education, including a higher degree in biology, as well as fifteen years teaching in classrooms, often racially diverse classrooms, I never heard a straightforward discussion about race and genetics.
Here, I should say, anthropology and the social sciences have done a marginally better job of disseminating their findings than biology and the natural sciences. Anthropologists have written about race, and if you study anthropology you may encounter some of these ideas. The predominant view of anthropology, however, does not tell the whole story, as I hope to show. Biologists have also written about race, and performed a great deal of research, but this work is not well known, and largely absent from academic and public discourse.
This is unfortunate because modern science offers a surprisingly clear view of race and genetics. Strong evidence supports the statements above, namely, that there are no significant genetic differences between people around the world, and that there are good reasons to believe that we will never find such differences. Despite the fact that we have known this for decades, racism continues to shape society with devastating consequences.
My goal, therefore, is to communicate the conclusions of modern science as clearly as possible, including background concepts, key findings, and supporting evidence. In this way, I hope to contribute to ongoing discussion of race and genetics.
I will make some attempt to explain why I think race and genetics is important. Some people believe that that there are no racial differences. All humans are created equal, and there is nothing more to say. Others believe racial differences are obvious, and it’s unnecessary to expend effort on further explanation. Personally, I think it’s impossible that any curious and thoughtful person could fail to find issues of race and racism engaging and critically important.
Since modern humans appeared, about 200,000 years ago, in East Africa, we have broken ourselves into groups, perceived differences between groups, and engaged in various forms of competition, cooperation, trade, exchange and war with other groups. The instinct to identify with one group and to view others with suspicion or hostility appear universal. There are plausible arguments that people who exhibited strong group identity, based on physical appearance, language, religion or any other criteria, were better able to compete for resources, defend and expand their territory and preserve their culture, compared to people who did not exhibit this behavior. Strong group identity may have conveyed a survival advantage upon our ancestors. In this sense, perhaps racism is natural. I think there is some truth to this, but I would counter that empathy toward and cooperation with other people would confer perhaps an equal survival advantage, and, for this reason, it is perhaps the tension between these two impulses—divisive and inclusive—that creates the play of humanity.
Picking up human history in the recent past, we confront the crusades, the colonial conquests, the ravages of imperialism, slavery, the great wars, the Holocaust, the partition of India, and so on—events based on beliefs in the supremacy of one group compared to another. These beliefs have persisted in the last fifty years, perhaps most tragically in the genocides in former Yugoslavia and Rwanda, and most obviously in ongoing conflicts in the Middle East, the violent conflict between the West and radical Islam, the growing tension between the West and the East, the looming prospect of a second Cold War, the emergence of what is perhaps a new civil rights movement in the United States, and the increasing arrival of and reaction against immigrants around the world, mainly refugees fleeing conflicts, themselves created by ethnic or religious tension.
This horrific legacy overwhelms the banal but no less keenly felt racism that exists in every country in the world, as evidenced by varying degrees and kinds of income inequality, opportunities for employment, social and political privilege, racial or ethnic mistreatment and religious intolerance. Once again, all of these are based on the belief that one group is superior to another, which is to say, on racist beliefs.
Short of the ravages of disease and the hardships imposed by the physical environment, it’s hard to think of anything that has been responsible for, and continues to cause, such suffering. I created this website to address these racist beliefs, and make sense of, and possible improve, the world.
Before moving on, there are several points I would like to make. First, I think it’s important to acknowledge that scientific discourse about race has a long and ignoble history of prejudice and personal and cultural bias. Our ideas about race and genetics have always reflected the sentiments and politics of the time. Recall that, not long ago, the scientific consensus was that there were differences between races. I promise to explain the findings of modern science, therefore, I must admit that these findings may be, in fact almost certainly are, in some way influenced by present political and cultural views. I mentioned that there is some reluctance to speak about race and genetic openly. Critics determined to find fault with modern science will undoubtedly interpret this as evidence that science is taking shelter behind a cloak of political correctness, to avoid telling the truth, which they presume to know. I offer that, instead, the present climate is an unfortunate though understandable response to the wrongs of the past, and a recognition of ongoing racial conflict. In any case, while it is certainly true that science, like all other human endeavors, is subjective, I do believe that science advances in small steps toward a better understanding of the natural world, informed by comprehensive data, with great explanatory power, and the potential to change our lives.
Second, I will make a necessary distinction between biology and culture. These are weighted terms but, broadly speaking, when we ask questions about race and genetics, we can seek biological explanations or cultural explanations. For example we can ask, “Are there variations in hair between groups of people from different parts of the world?” Obviously, some differences, such as natural hair color and type are determined by biology, while other differences, such as the way that hair is decorated or dyed, are determined by culture. In a similar fashion, we can ask, “Are there variations in intelligence between groups of people around the world?” I have already indicated that, insofar as science can determine, there are no such differences, however, particular aptitudes commonly associated with intelligence, such as reading level or mathematical ability, are strongly linked to education, income, infrastructure, and so forth, which is to say, to culture.
Culture is one of the great forces that shapes human civilization. The acquisition of language, art, music, religion, complex tools, shelters, clothing and jewelry, and the potential to transmit these artifacts horizontally through populations and vertically through time, is one of the landmarks of human evolution. Some scientists believe that the acquisition of culture marked a shift from an era governed mainly by genetic evolution to an era marked primarily by cultural evolution. The answers to many questions about human diversity are cultural. Indeed, if what I have suggested is true—once again, that there are no significant biological differences between groups of people—then variation must be largely cultural.
I will focus on biology. I was trained as a biologist, not as an anthropologist or sociologist, and I am not skilled enough to parse the layers of meaning embedded within our society, to say nothing of comparing societies around the world. In any case, culture has the great advantage of being malleable, at least over timescales that are meaningful to us, compared to slow working changes of genes.
Last, it will probably be helpful if I explain the kinds of questions that I will address. When we talk about race and genetics, we are usually not overly concerned with things like eye shape or hair style. Instead, we want to know if there are racial differences in intelligence, aggression, love, violence, ambition, sexual prowess, criminality, empathy, religious faith and health. I call these fundamental characteristics, to distinguish them from superficial physical characteristics. The distinction between fundamental characteristics and superficial physical characteristics is an important part of my arguments.
Species and sub species
When we ask, “Are there differences between races?” or, “Are there races?” what exactly do we mean? The word race appears to be derived from the 16th century Middle French race, “people of common descent,” related to the Italian razza, and the Spanish and Portuguese raza, which mean, “Breed, lineage, or family.” Dictionaries variously describe race as 1) a breeding stock of animals, 2) a family, tribe, people, or nation belonging to the same stock, 3) a category of humankind that shares certain distinctive physical traits, and 4) a class or kind of people unified by interests, habits, or characteristics. Synonyms of race include: ethnic group, population, community, line, lineage, family, breed, people, species, nation, stock, tribe, kin, and clan. How can we make sense of these multiple overlapping definitions, influenced as they are by history and culture? Some words, such as “breed” or “stock” are offensive, as they suggest close infinity to domesticated animals. Other words, such as “tribe” or “clan” invoke primitive people. Still others, such as “community” or “family” are so vague that they prove essentially meaningless. We need a more specific word for race.
One candidate is the biological term species. Most people have at least some familiarity with Linnaean classification, the system commonly used to organize and name living things. Linnaean classification is based on a series of hierarchical categories or groups. Larger groups contain smaller groups, which contain smaller groups, and so on, like a series of nested boxes. The largest group is all life on Earth. The smallest unit is each individual life form.
These categories have evolved over time, and are subject to ongoing modification. They are, from largest to smallest: Domain, Kingdom, Phylum, Class, Order, Family, Genus and species. The last two categories, Genus and species, are written as such, Genus capitalized and species not, both italicized, most often provided together, a convention called binomial nomenclature, which assigns all living things a two part name. Humans are therefore Homo sapiens, although even this is controversial, as I will discuss below.
(I should say that Carl Linnnaeus (1707-1778), the Swedish biologist who invented this classification system, widely regarded as the “father of modern taxonomy,” proposed one of the first very offensive classification system for humans. As I mentioned above, early attempts in this field reflect poorly on scientists and the scientific process, and should be viewed as expressions of cultural beliefs at the time, and not evidence of underlying biological differences.)
It’s actually more difficult that one might think to define a species. I can offer the following: a species is a group of living things that breeds successfully under normal conditions in the wild. This might seem unnecessarily complex—why not simply say that a species is a single living thing? In fact, each part of the definition is important. Consider the following domesticated draft animals: the horse, the donkey and the mule. Horses can all breed together successfully, producing many familiar varieties, from sleek Arabians to imposing Clydesdales. Likewise, donkeys can all breed together successfully. In contrast, while horses and donkeys can breed with one another, the resulting animal, a mule, is infertile. Therefore, the pairing of horses and donkeys is not successful in a biological sense, and the two equids remain separate.
A mule is considered a hybrid, common among plants, less so among animals, although various animal hybrids do exist: the liger (lion and tiger), the zonkey (zebra and donkey), or the wolphin (whale and dolphin). These creatures seem outlandish tricks of nature, but they do exist, and they serve to illustrate an important point: while some creatures can breed successfully, they usually do not in the wild. One example that comes to mind involves large felines, such as leopards and jaguars. Leopards are confined to Africa and Eurasia, Jaguars to South and Central America. The two never meet under natural circumstances, however, if they did, they could most likely breed successfully. This demonstrates the importance of geographic barriers, and difficulty of clearly defining species.
Obviously, all humans belong to the same species. Humans from every corner of the planet can and do breed with one another, overcoming formidable geographic and cultural barriers. Inuit people can have children with Javanese islanders, natives of Patagonia can produce families with Koreans or Malaysians or Madagascarians and so forth. This serves as indelible evidence that we are one species
Since the appearance of hominins, upright walking, large brained primates, about six million years ago, there have been times when multiple hominin species appear to have lived together, presumably with overlapping territories, and associated conflicts and couplings. For example, about two million years ago, there may have been at least five different hominins roaming Africa. Whether or not these creatures reproduced is not clear, although we may reasonably infer, from studying their remains, that they were probably as different from one another as the remaining great apes—gorillas, orangutans, chimpanzees, and bonobos, which do not breed successfully.
It’s fascinating to imagine what it would be like if one or several different hominin species existed today. I often show my students an artist’s rendering of, for example, Homo habilis, the so called “handy man,” one of the first tool-making hominins, who, despite his upright stance and manual dexterity, was likely a squat, long-armed, low-browed creature, covered in hair. I ask, “Would you have sex with this person?” This question has important implications. We believe that Homo habilis likely did not breed with Homo rudolfensis, Homo egaster, Paranthropus boisei, Paranthropus robustus, Parathropus aethiopicus, or any of the other hominins that lived at the same time in the same place. However, recent genetic evidence suggests that humans did breed with Neanderthals, and, perhaps, with another group, Denisovans. If true, some scientists argue, humans, Neanderthals and Denisovans can not have been different species, and should be properly classified as Homo sapiens sapiens, Homo sapiens neanderthalensis, and Homo sapiens denisova.
We thus arrive at the important idea of sub-species. At the outset, I would like to say that the word “sub” meaning below, and thus inferior, would appear dangerous in relation to any discussion of race and genetics. However, following the logic of taxonomy, sub-species are simply smaller groups nested within larger groups, and the term is broadly used without any pejorative connotations. In biology, the words “race” and “sub-species” are synonymous. Therefore, when we ask, “Are there human races?” we are really asking, “Are there human sub-species?”
Sub-species are populations that have become sufficiently different, in terms of their body shape, size, coloration or any other feature, that they deserve independent classification. Theoretically, they could reproduce successfully, but they do now. The division of species into sub-species usually occurs because of some kind of barrier, which could be as simple as a river cutting through a meadow, or as dramatic as a mountain range bisecting a continent. The separation need not be physical. Imagine one group of animals feeding during the day and another feeding at the night, or one group of trees dispersing their seeds in spring and another in summer. The important point is that the chance of breeding, or gene flow, between the two populations is reduced, and so they begin to evolve along distinct paths. Sub-species, you could say, are on their way to becoming species, although they have not yet accumulated enough genetic differences to be classified as such.
Traditional classification
How different must one group be from another group to constitute a sub-species? As you can imagine, this is the subject of much controversy. Entire careers have been staked upon the precise taxonomic definition of a particular plant, beetle or bird. Traditionally, scientists used physical features to classify sub-species. There are several problems with this method, and I think it’s worth talking about them in detail, because they illustrate the difficulty of defining human races.
First, many physical characteristics change gradually from place to place. I call this the gradual change problem. Consider skin color. Despite the fact that skin color is so often used to define human races, to the extent that races are often described by their color (black, white, yellow, red) pigmentation changes gradually. People from equatorial regions and lower latitudes generally have darker skin, while people from higher latitudes generally have lighter skin, but between extremes there is nearly infinite variation, one shade blending seamlessly into the next, without clear distinctions between them. The scientific term for this kind of change is clinal variation. Skin color changes along a cline. Therefore, classifying individuals based on skin color is difficult. It’s relatively easy to distinguish native people from Scandinavia and Australia, but nearly impossible to do so for inhabitants of Mexico and Guatemala, or any two adjoining regions, based purely on skin color. The same is true of most other physical characteristics. Skin color is an important topic that I will discuss in more detail later.
Second, many characteristics do not correlate with each other in predictable ways. I call this the correlation problem. There are several examples among humans. The epicanthic fold, an extension of the upper eyelid that covers the inner corner of the eye, is usually associated with Asians. However, other groups of people, such as the San of South Africa, also have the epicanthic fold. Blonde hair and blue eyes are associated with Northern Europeans, although both features can be found among people from India and Melanesia. Straight hair is associated with several groups of people, but not with Australian aborigines, although aborigines with straight hair exist. I don’t want to overstate these exceptions, but simply note that many physical features which we think are found together are sometimes not, features that we think occur independently sometimes coexist, and this makes classification difficult.
Third, because many characteristics change gradually from place to place, and do not correlate with each other in predictable ways, defining meaningful groups is troublesome. I call this the group problem. For example, if you set out to define two broad groups, Europeans and Asians, what feature or features would you use? If you chose skin color, you would find that pigmentation changes along a cline from north to south. If you chose hair color, you would find that black hair is widely distributed throughout Eurasia. If you chose the presence or absence of the epicanthic fold, you would find differences between western Europe and eastern Asia, but you would also find that the eyes of native Greenlanders, for example, bear more resemblance to those of Mongolians than those of their closer neighbors in Scandinavia. Of course, you could define any number of groups, but this is precisely the problem. Groups often have more to do with the criteria you select, rather than features of populations.
Finally, we often set boundaries or thresholds to classify living things. These boundaries may be arbitrary. Imagine the implications if you decided that a particular shade of skin defined black or white, or a particular shape of eye defined Asian or not Asian. I call this the threshold problem, and it bedevils many classification systems.
More recently, scientists have employed genetic analysis. Allowing that genetic analysis is associated with its own problems, it offers several advantages. After all, the division of species into sub-species is essentially about gene flow, and so it makes sense that techniques that study genes directly provide the most satisfactory answers.
Genetic analysis
Scientists use variety of genetic techniques to classify species and sub-species. One method is the measurement of genetic distance. Genetic distance describes how far the genomes of two organisms have diverged from another because of limited gene flow between them. It is usually expressed numerically as the fixation index, abbreviated FST, ranging in value from 0 to 1. A FST of 0 describes a population of freely interbreeding individuals, while a FST of 1 describes two populations that are completely separated from one another. In most cases FST is neither 0 nor 1 but varies somewhere in between.
Despite the apparent precision of this technique, FST values change depending on which genetic markers are selected, and which statistical analysis are applied. Moreover, there are no universal guidelines about where exactly where to set the threshold to determine sub species. Rather, the threshold to determine sub species changes among different groups, such as plants, birds, reptiles, amphibians and so on. Nonetheless, this technique has been applied successfully across a broad range of species in varying contexts, thus we can be reasonably confident about its validity. What, then, does genetic analysis tell us about human diversity?
Here we arrive at the first outstanding and critically important finding of modern science. The generally accepted threshold to define sub species of large mammals is 0.25. If the FST of two large mammal populations is below 0.25 they are not different sub-species, and if the FST is above 0.25 they are different sub-species. Genetics analysis shows that the global average human FST varies between 0.05 and 0.15. This number falls well below the threshold to classify sub-species of large mammals, and so we conclude that there are no human sub-species, and in the classical biological sense no human races. If more people understood this fact, one feels, the world would be considerably improved.
Because this point is so important–the human global FST falls below the threshold to classify sub-species of large mammals and so there are no human races–it’s worth spending some time examining these concepts more closely.
Figure 1 shows a range of FST values from 0 to 1. The dark blue line represents an FST of 0.25, the threshold to classify sub-species of large mammals. Because FST values are subject to some uncertainty, it is probably more accurate to define a range within which it might be reasonable to assume that two populations are sub-species. This range, from 0.2 to 0.3, is show in light blue. The average global human genetic distance is show in light green. As you can see, the light green band falls well below the light blue band.
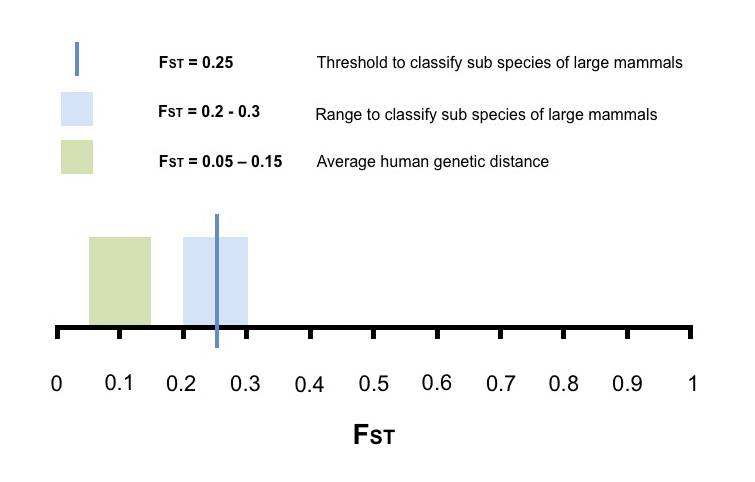
Figure 1. Fst threshold to classify sup species of large mammals, and average global human genetic distance.
Figure 2 shows human FST values obtained for a large number of genetic markers. I will discuss genetics in more detail below, however, briefly, genetic markers are small pieces of DNA that can be examined and compared between individuals. In this case, the genetic makers are single nucleotide polymorphisms (SNPs) that vary at at one specific place or nucleotide base. American biologist Sean Myles and his colleagues examined 2750 SNPs from 927 individuals from 53 populations around the world. The Y axis shows the numbers of SNPs, and the X axis shows the corresponding FST .
There are several important conclusions that we can draw from these data. First, we can see that for the great majority genetic markers the FST is very low, in most cases far below the threshold to classify sub-species of large mammals. The greatest numbers of genetic markers have an average FST of perhaps 0.05, which confirms previous studies of human genetic distance. Second, we can see that FST values change depending on the number of genetic markers included in the analysis, as well as the particular genetic markers selected. If we studied only one SNP, or ten, or twenty, we might reach false conclusions about human genetic diversity. For example, if we choose an SNP with a very low FST, such as 0.01, we might conclude that all humans are joined in a single freely-breeding population with absolutely no barriers to gene flow. This is not the case; while all humans can breed together successfully, there have always been and remain some barriers to gene flow, both geographic and cultural. Likewise, if we choose SNPs which showed very high FST, such as 0.6, we might conclude that different populations should be classified as sub-species or races. A complete picture of humans diversity emerges only when we examine a great number and variety of genetic markers.
Some readers will inevitably claim that the few SNPs with a high FST prove that human races exist. I will respond in more detail later, but for the moment, I would simply like to point out that SNPs are randomly pieces of DNA which most likely bear no relation to recognizable characteristics, not least the characteristics that we are concerned about when we talk about racial differences. If pressed, I would say that SNPs which show a high FST might be related to genes responsible for obvious physical differences between people from around the world, but I suspect that, instead, they reflect our diverse origins and evolutionary histories. DNA is an admixture of fragments that came together from all over the world at different times. It should come as no surprise, therefore, that some locations on DNA show significant diversity.
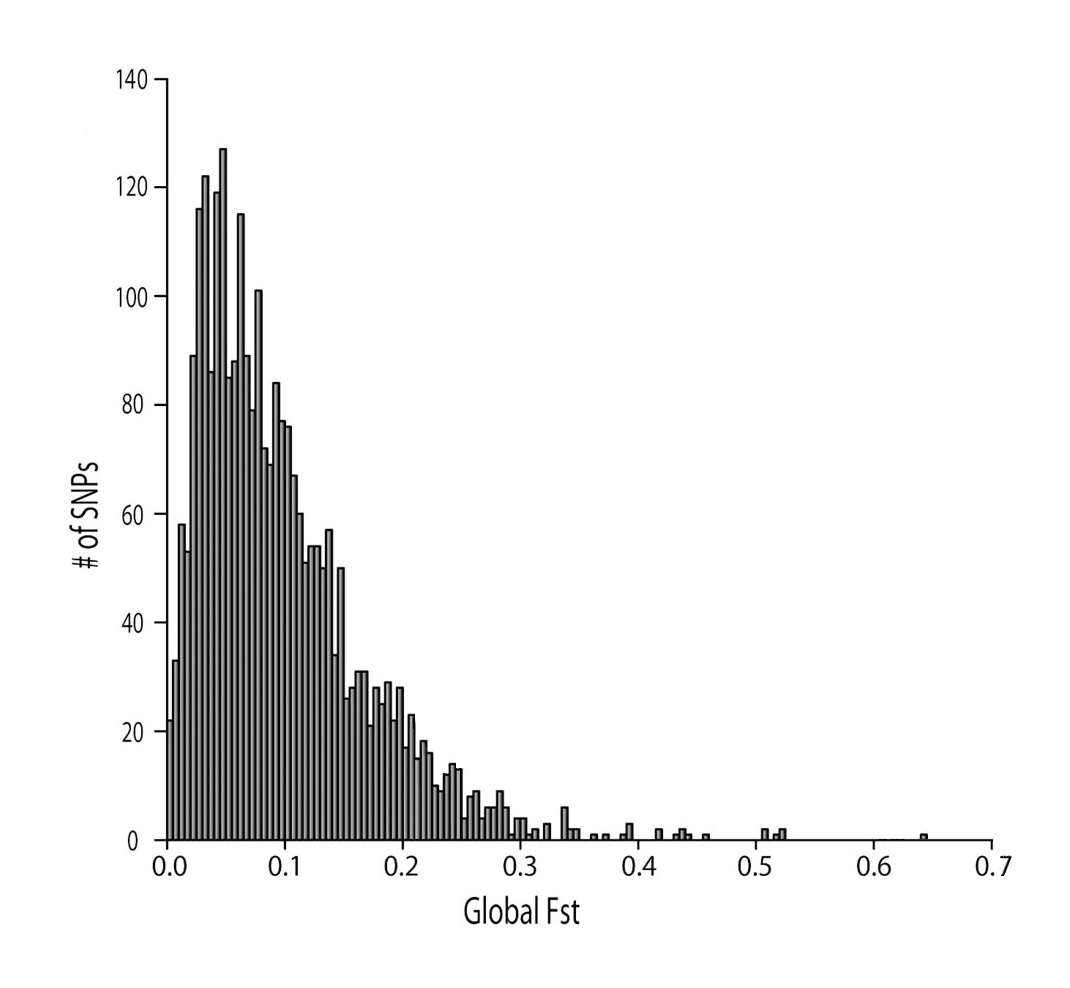 Figure 2. Global human FST distribution of multiple genetic markers. Adapted from Myles et al. 2008.
Figure 2. Global human FST distribution of multiple genetic markers. Adapted from Myles et al. 2008.
Figure 3 shows the genetic distance of humans in comparison to other large mammals. The Y axis shows average FST values, and the X axis shows a variety of large mammals. The height of the blue bars corresponds to genetic distance. Again, human genetic distance is very low, below the FST of many other large mammals and primates, including chimpanzees, our closest ancestors. This places human genetic diversity in context, and raises an important question: “Why is human genetic distance so low?” The answer to this question is the subject of our next discussion.
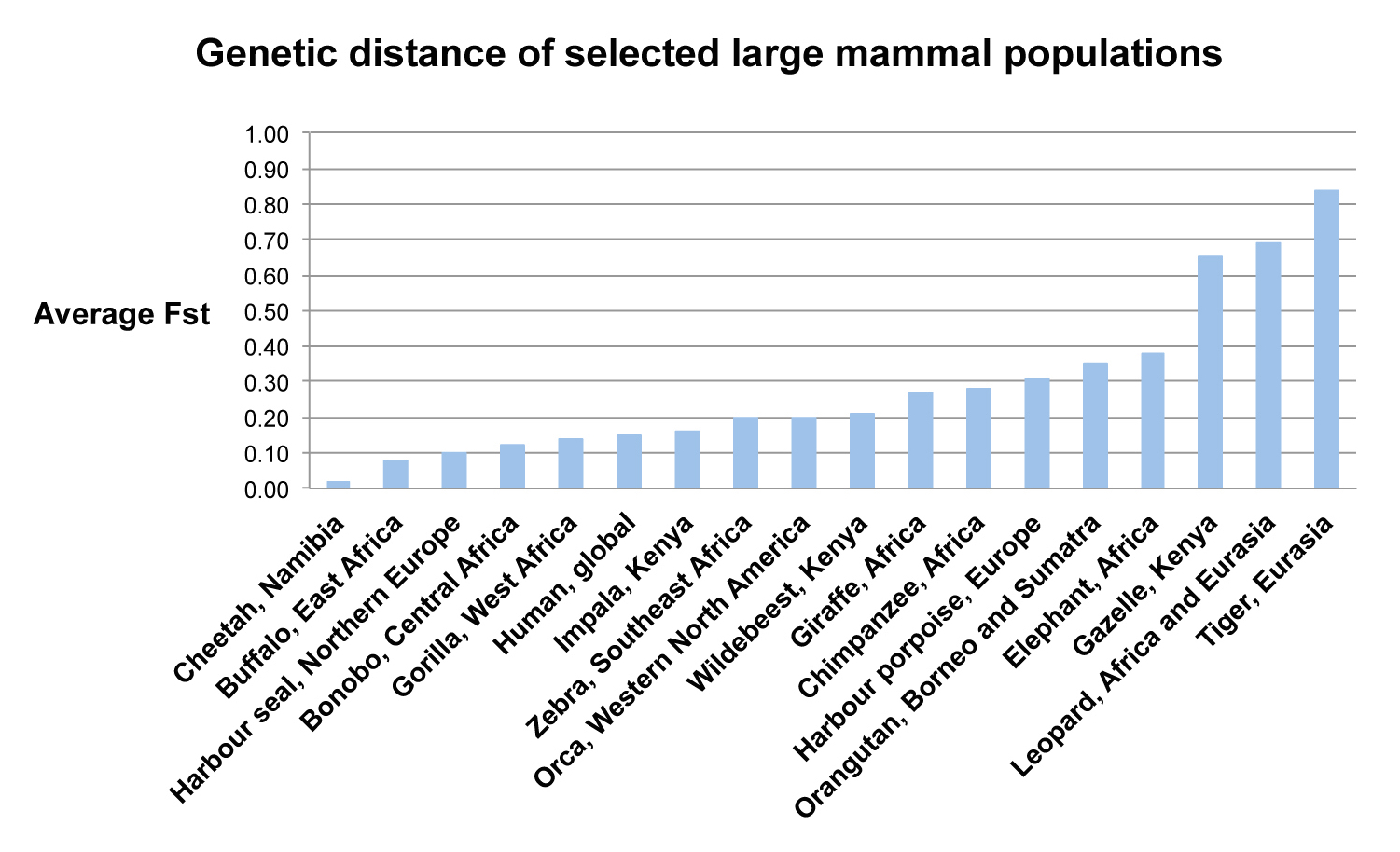
Figure 3. Genetic distance of selected large mammal populations. Values were obtained from the following sources: Cheetah, Marker et al. 2008; Harbour seal, Andersen et al. 2010; Bonobo, Fischer et al. 2006; Gorilla, Fischer et al. 2006; Human, Barbujani et al. 1997; Zebra, Lorenzen et al. 2008; Orca, Barret-Lennard and Ellis 2001; Giraffe, Brown et al. 2007; Chimpanzee, Fischer et al. 2006; Harbour porpoise, Fontaine et al. 2007; Orangutan, Fischer et al. 2006; Leopard, Uphyrkina et al. 2001; Tiger, Luo et al. 2004; Al. other values from Templeton 1998.
How we became so similar
In addition to providing a clear answer about whether or not there are human races—there are none—modern science can also explain why humans are so similar. Some researchers believe that, approximately 75,000 years ago, before humans left Africa in large numbers and dispersed throughout the world, there was a cataclysmic environmental event known as the Toba catastrophe. During the Toba catastrophe, an enormous volcanic eruption occurred on the island of present day Sumatra. Ash spread throughout the atmosphere, blocking solar radiation, and there was a pronounced drop in temperature, known as a volcanic winter, followed by a general cool period, which may have lasted more than 1,000 years. The human population, by some estimates, was reduced to as few as 10,000 individuals. All humans alive today, close to 7.5 billion people, are descendants of this small ancestral group of survivors. It stands to reason that all humans are very similar genetically.
This phenomena, by which a large population is reduced in size and diversity, is known as a genetic bottleneck. Genetic bottlenecks are caused by a number of factors, including environmental change, natural disasters, competition with other species, disease or migration. The original population passes through the narrow “neck of the bottle” and only a small sample of the original population remains. Following the laws of probability, genetic diversity is reduced. To make this point clear, imagine a bag containing a great number of marbles of different colors, shapes, and sizes. If you reached into the bag and withdrew a handful of marbles, in all likelihood, the small sample you selected would not reflect the same diversity as the original. This process is shown in the figure below.
 Figure 4. Genetic bottleneck. Simon EJ, Dickey JL, Reece JB, 2013. Campbell Essential Biology, Fifth Edition. Pearson Education, Inc., Upper Saddle River, NJ.
Figure 4. Genetic bottleneck. Simon EJ, Dickey JL, Reece JB, 2013. Campbell Essential Biology, Fifth Edition. Pearson Education, Inc., Upper Saddle River, NJ.
The Toba catastrophe is one example of a human genetic bottleneck. Our subsequent migration from Africa is another. These events explain our genetic similarity.
As an aside, bottlenecks generally have the effect of reducing genetic diversity. However, similar process, acting in a slightly different ways, can also contribute to differences between populations. I will discuss these processes below.
The next step
Many people stop at this point. The problems with traditional classification make it difficult or impossible to separate humans into meaningful groups. Genetic analysis shows that all humans are extremely similar. There is no scientific basis for sub-species, and so there are no human races.
This view is shared by many anthropologists. In academic circles, you often hear, “There is no such thing as human race,” or, “Human races do not exist.” I agree with these statements wholeheartedly, in the sense that conventional racial categories, such as black and white, are meaningless, and do not adequately reflect human diversity.
At the same time, I don’t think this is a satisfactory explanation of race and genetics. It’s not satisfying to my students, perhaps the most discerning and astute arbiters of truth, and it’s not satisfying to the great majority of people.
After all, humans from different parts of the world clearly look different, and clearly act different, therefore, it’s only natural to believe that some meaningful genetic variation exists. Likewise, it’s not sufficient to explain human variation entirely as a product of culture. As I pointed out, while culture is one of the great driving forces for human enterprise, it can not explain variation in physical characteristics which are obviously genetic, or have a strong genetic component, such as skin color, height, facial features and so on. These differences must be related to genetics, and any explanation that fails to account for them is incomplete.
The preeminent British biologist, Richard Dawkins, treated this subject with considerable clarity and characteristic intellectual honesty in his remarkable book, The Ancestor’s Tale, A Pilgrimage to the Dawn of Evolution. He explained the idea of inter-observer correlation, roughly that, when faced with complex problems, where clear solutions are not immediately obvious, useful information may be obtained if multiple observers reach similar conclusions. More generally, it makes sense to pay attention if many people reach the same conclusion. He pointed out that human variation shows a high-degree of inter-observer correlation, and suggested the following experiment:
“Suppose we took standard full-face photographs of 20 randomly chosen natives of each of the following countries: Japan, Uganda, Iceland, Sri Lanka, Papua new Guinea and Egypt. If we presented all 120 people with all 120 photographs, my guess is that every single one of them would achieve 100 per cent success rates in sorting them into six different categories. What is more, if we told them the names of the six countries involved, all 120 subjects, if they were reasonably well-educated, would correctly assign all 120 photographs to the correct countries.”
To be clear, Dawkins does not extend this reasoning to suggest that there are human races, nor that human differences involve anything other than superficial physical characteristics. His point is that differences between groups of native people around the world show high inter-observer correlation, and that we must acknowledge and explain this fact if we are to understand race and genetics. I would therefore like to turn my attention to human genetic variation, what it means, and, more important, what it does not mean
Genetic variation
The American biologist, Richard Lewontin, published one of the most important studies about human variation, The Apportionment of Human Diversity. Lewontin compared seventeen genetic markers from seven groups: Caucasians, Africans, Asians, South Asians, American Indians, New Guineans, and Australian Aborigines. He found that fully 85 percent of genetic variation was shared by all populations, while only 15 percent was unique to any one population. He concluded,
“It is clear that our perception of relatively large differences between human races and subgroups, as compared to the variation within these groups, is indeed a biased perception and that, based on randomly chosen genetic differences, human races and populations are remarkably similar to each other, with the largest part by far of human variation being accounted for by the differences between individuals. Human racial classification is of no social value and is positively destructive of social and human relations. Since such racial classification is now seen to be of virtually no genetic or taxonomic significance either, no justification can be offered for its continuance.”
There were several important limitations to Lewontin’s study, including the small number of genetic markers, the difficulty classifying humans into groups that have biological significance, and the fact that diverse groups, such as all Africans or all Asians, were considered together. Nonetheless, his results have been confirmed by numerous subsequent and more rigorous studies. Researchers have repeatedly found that 75 to 85 percent of human genetic variation is shared between groups, while 25 to 15 percent is unique to any particular group.
In many ways, this confirms what I explained above, namely, that we are far more similar than we are different. It is remarkable that scientists have known this for nearly fifty years, and yet it has not become common knowledge. Part of the problem is that it’s not easy to understand exactly what Lewontin’s numbers mean. What is genetic variation, and how can it be partitioned between and within groups?
To understand Lewontin’s work, we have to review the basic science of genetics. Hereditary information, as most people know, is stored in an elegant molecule called deoxyribonucleic acid, or DNA. The structure of DNA is perhaps most easily understood as a ladder twisted into a helix. The long uprights, or backbones, of the DNA ladder, are composed of alternating ribose and phosphate molecules, while the short repeating units, or rungs, are composed of pairs of molecules called nucleotide bases. There are four nucleotide bases: adenine, thymine, cytosine, and guanine, often abbreviated, A, T, C, and G, that follow particular pairing rules, to preserve the symmetry of the composite structure.
Each half of DNA, like one side of a zipper, therefore contains one long backbone, and a series of single nucleotide bases, arranged in a constantly varying sequence. It is this sequence of bases that stores information. In much the same way that long sequences of 0s and 1s encode digital information, long sequences of A, T, C, and G encode biological information. Specifically, the sequences of nucleotide bases contain the instructions to make proteins, the principal molecules that create and regulate living things.
The figure below shows a simple representation of DNA. On the left you can see the double helix, in the middle you can see an expanded view, and on the right you can see nucleotide base pairing.
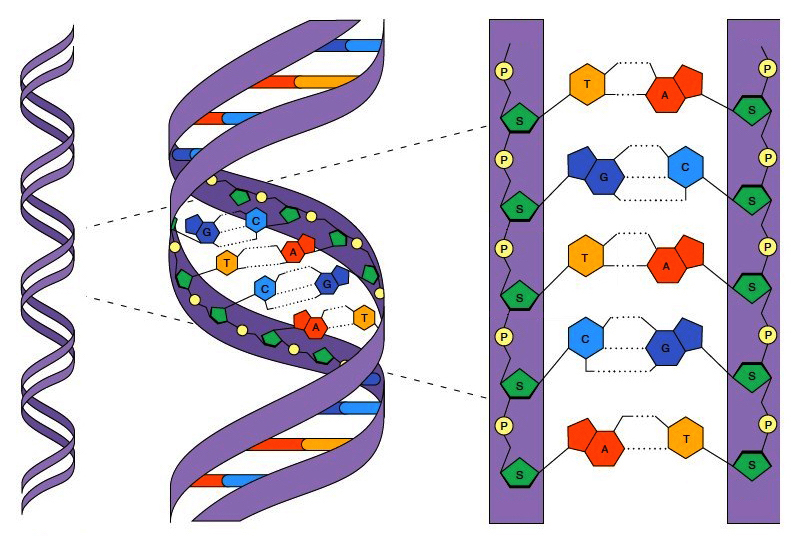 Figure 5. The structure of DNA. Genetic bottleneck. Simon EJ, Dickey JL, Reece JB, 2013. Campbell Essential Biology, Fifth Edition. Pearson Education, Inc., Upper Saddle River, NJ.
Figure 5. The structure of DNA. Genetic bottleneck. Simon EJ, Dickey JL, Reece JB, 2013. Campbell Essential Biology, Fifth Edition. Pearson Education, Inc., Upper Saddle River, NJ.
It is absolutely astonishing to consider that every person has unique sequences of nucleotide bases in their DNA, which create unique combinations of proteins, which create unique individuals. The cells in my body contain DNA molecules with particular sequences of nucleotide bases, which contain the information to make particular proteins, which makes me me. The cells in your body contain other DNA molecules with other sequences of nucleotide bases, which contain the information to make other or slightly different proteins, which makes you you. And so on, for every person on Earth.
It is not the case, however, that the DNA of any two people is completely different from beginning to end. The overwhelming majority of DNA is shared. Therefore, if we compare the DNA of two people, we find long sequences of bases that are identical, and only a small number of bases that are different. Scientists estimate the DNA sequences of any two humans vary by only one half of one percent. Nonetheless, this is the variation that creates differences between people, and this is the variation that biologists study.
Genetic markers, as explained above, are small pieces of DNA of varying length, or in the case of SNPs, single nucleotide bases. By comparing the sequences of genetic markers, scientists can quantify genetic variation, or the extent to which individuals vary from one another. When scientists say that two groups share 85 percent of genetic variation, or that 15 percent of genetic variation is unique to any particular group, they are referring to this kind of variation.
The figure below shows the genetic variation between two native populations from different continents. Each population consists of one hundred people. The letters inside the boxes represent variations of a genetic marker composed of four nucleotide bases. If you compare the two populations, you can see that nearly all of the sequences are shared by both groups (blue boxes), while only a small portion of the sequences are unique to one group or another (green boxes). By counting the boxes you can quantify the variation within and between the populations: 85 boxes or 85 percent of genetic variation is shared by both populations, while 15 boxes or 15 percent of genetic variation is unique to each population. This is what Lewontin discovered about human genetic variation.
Another way to interpret this figure is to consider what I call the “island scenario.” Imagine that each population lived on a different island. If one population went extinct, because of some kind of environmental catastrophe, such as a volcanic eruption, earthquake, tsunami, famine or disease, the great majority of genetic variation would be preserved in the remaining population.
This example is fictional. In reality, the numbers of individuals and genetic markers would not be so straightforward to analyze. However, I have found that the figure helps people understand the nature of genetic variation.
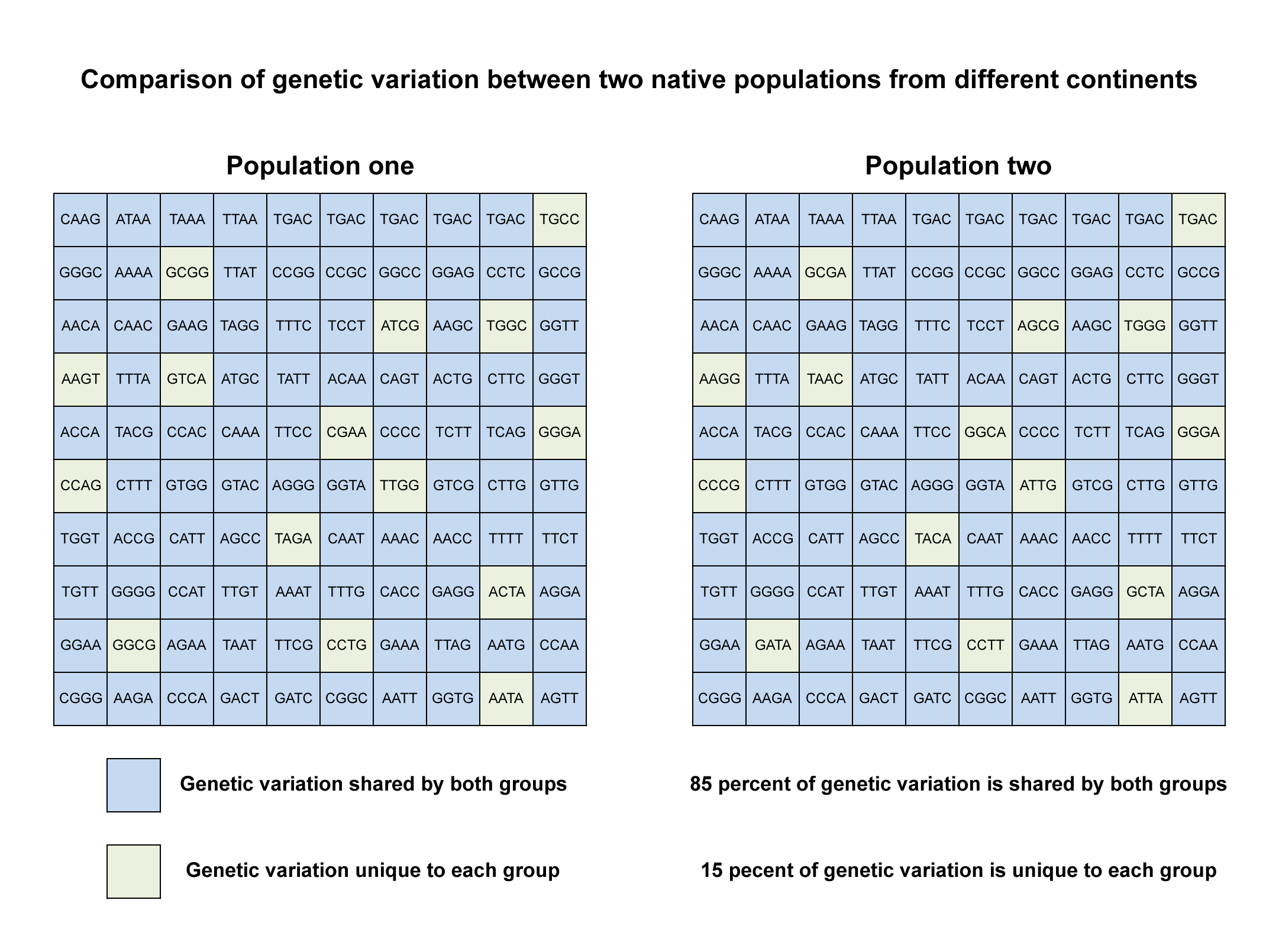 Figure 6. Comparison of genetic variation between two native populations from different continents.
Figure 6. Comparison of genetic variation between two native populations from different continents.
Lewontin’s findings have been confirmed repeatedly and generally accepted by the scientific community, but several prominent figures have criticized his work. Perhaps the most significant response came from another British biologist, Anthony William Fairbank Edwards, who published a paper entitled, Human Genetic Diversity: Lewontin’s Fallacy. Edwards claims that Lewontin’s work ignores the fact that certain variables are linked or associated, and that these associations are responsible for significant differences between groups of people around the world:
“In popular articles that play down the genetic differences among human populations, it is often stated that about 85% of the total genetic variation is due to individual differences within populations and only 15% to differences between populations or ethnic groups. It has therefore been proposed that the division of Homo sapiens into these groups is not justified by the genetic data. This conclusion, due to R.C. Lewontin in 1972, is unwarranted because the argument ignores the fact that most of the information that distinguishes populations is hidden in the correlation structure of the data and not simply in the variation of the individual factors.”
Insofar as I can follow Edward’s general argument, if not the complex mathematical analyses that comprise the bulk of his paper, I am inclined to agree. Considering that human groups show a high-degree of inter-observer correlation, there must be some meaningful genetic variation. Dawkins, in the The Ancestor’s Tale, confirms this view:
“It is genuinely true that, if you measure the total variation in the human species and then partition it into a between-race component and a within-race component, the between-race component is a very small fraction of the total. Most of the variation among humans can be found within races as well as between them. Only a small admixture of extra variation distinguishes races from each other. That is all correct. What is not correct is the inference that race is somehow a meaningless concept.”
And,
“We can all happily agree that human racial classification is of no social value and is positively destructive of social and human relations. That is one reason why I object to ticking boxes in forms and why I object to positive discrimination in job selection. But that doesn’t mean that race is of ‘virtually no genetic of taxonomic significance.’ This is Edward’s point, and he reasons as follows. However small the racial partition of the total variation may be, if such racial characteristics as there are highly correlated with other racial characteristics, they are by definition informative, and therefore of taxonomic significance.”
Dawkins takes great care to explain that he does not believe there are fundamental differences between groups. Rather, he believes that differences are limited to superficial physical characteristics. Edwards is not so direct. He suggests that there are significant differences between groups, but he does not explain what he thinks these differences mean. Subjects such as race and genetics deserve greater clarity. If Edwards articulated his views about racial differences, his arguments would have greater power. That said, he does make a very important point:
“A proper analysis of human data reveals a substantial amount of information about genetic differences. What use, if any, one makes of it is quite another matter. But it is a dangerous mistake to premise the moral equality of human beings on biological similarity because dissimilarity, once revealed, then becomes an argument for moral inequality.”
Edwards highlights the moral dilemmas posed by scientific research about race and genetics. As before, some people will inevitably focus on the differences, however small, between groups. Therefore, I think it’s important to stop and place these differences in context. 1) Any differences between groups exist against a backdrop of overwhelming genetic similarity. 2) We are all same species, and there is no biological basis to classify humans as sub-species or races. 3) The differences that do exist are limited to superficial physical characteristics. 4) There is no evidence that there are fundamental differences. 5) We have very good reasons to believe that we will never find such differences (of which more later).
As long as we bear these points in mind, I think it’s possible, even important, to recognize that regular patterns of genetic variation do exist between groups. These differences play a large part in creating our perceptions of human races, and it seems appropriate to examine them more closely. We can therefore ask, “What are the observable patterns of human variation?” and, “How were these patterns created?”
Modern research
Studies of genetic variation gain power from numbers. If more genetic markers are examined, from more individuals, with more sophisticated techniques, we can be relatively more confident about the results, compared to studies that examine fewer genetic markers, from fewer individuals, with less sophisticated techniques. As I mentioned, Lewontin’s original study examined seventeen genetic markers, from seven groups, with common tools available at that time. More recent studies have examined considerably more genetic markers, from more groups, with far more powerful tools, including computers that can perform complex statistical analyses that would have been impossible to conduct in the past. The picture of genetic variation that has emerged from these studies is illuminating.
The figures below show results from several recent studies about human genetic variation. While the figures appear quite different in content and form, they tell similar stories. They show the genetic distance, or genetic variation, between native populations around the world. The simplest way to interpret these figures is to think about genetic distance in terms of geometry: the genetic distance between two populations is simply the Euclidean distance measured between two points.
In the first figure, the greatest genetic distance is between San Bushmen of South Africa and Australian Aborigines. In the second figure, the greatest genetic distance is between San Bushmen and natives of Papua New Guinea. In the third figure, the greatest genetic distance is between Xhosa of South Africa and the natives of the Amazon basin.
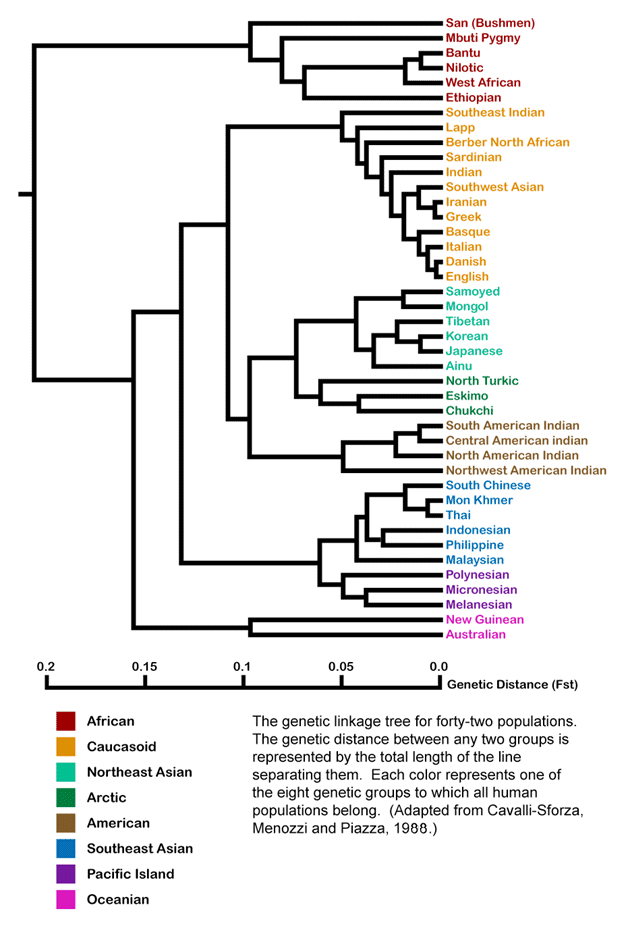 Figure 7. Human genetic variation. Cavalli-Sforza et al. 1988.
Figure 7. Human genetic variation. Cavalli-Sforza et al. 1988.
 Figure 6. Human genetic variation. Li et al. 2008.
Figure 6. Human genetic variation. Li et al. 2008.

Figure 8. Human genetic variation. Alvarado-Hernandez et al. 2011.
Which two groups are separated by the greatest distance? The San Bushmen and Australian Aborigines? The Xhosa and Amerindians? The answer is less important than the overall lesson: the greatest genetic distances are found between groups that are separated by the longest migration paths, and by the most formidable natural barriers, such as deserts, oceans and mountain ranges.
We can see this in the results of another study. The figure below shows people from South Africa, the Americas and Oceania arranged in a triangle, separated by the greatest distances, representing high genetic variation. People from Europe, the Middle East, and South and East Asia are clustered together in the middle, separated by less distance, reflecting low genetic variation.
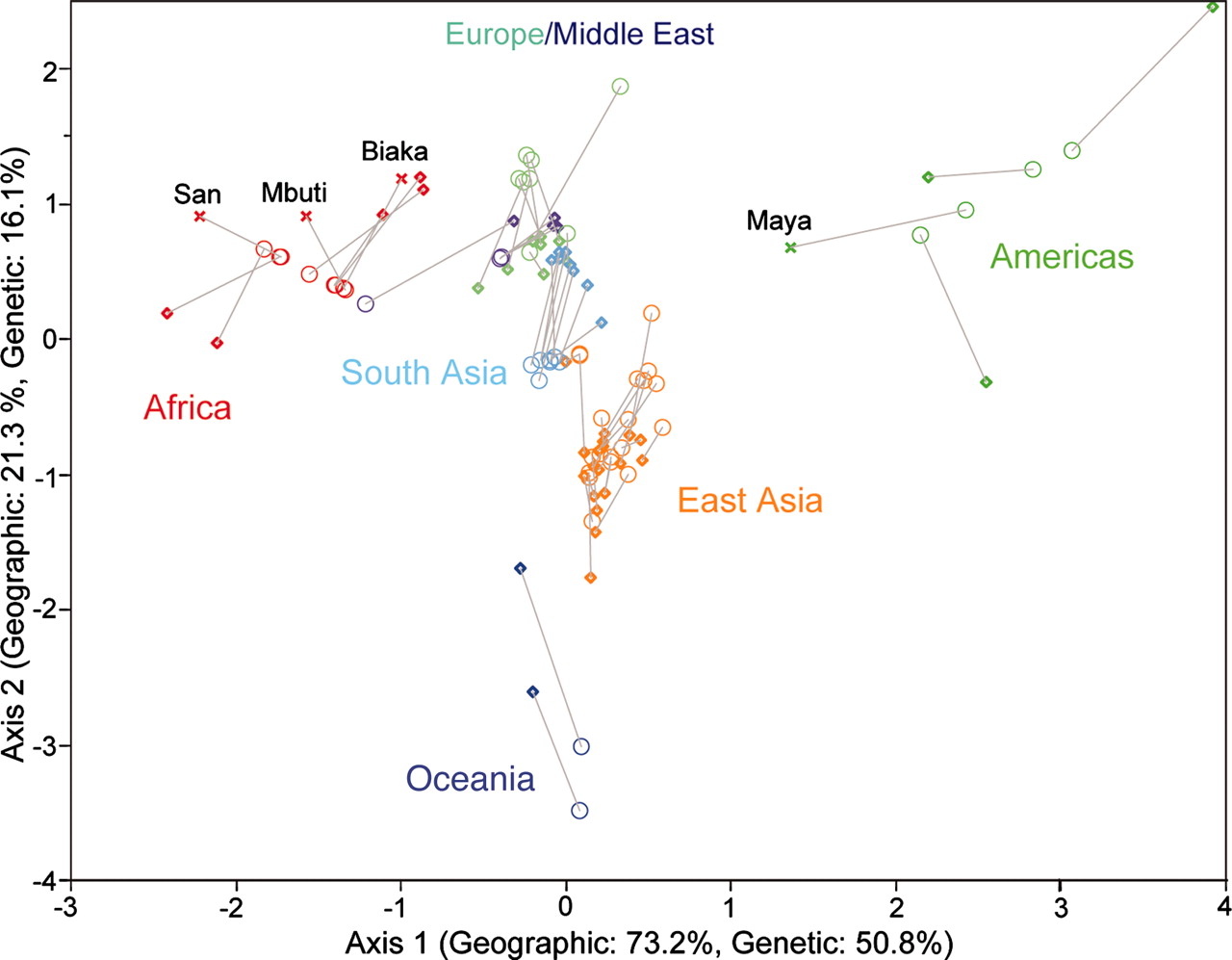
Figure 9. Human genetic variation. Ramachandran et al. 2005
This interpretation is satisfying because it supports, and is supported by, what we know about human migration. To appreciate this concordance, we need to consider our history.
About 65,000 years ago, following the Toba catastrophe, humans left Africa in large numbers, and began to disperse around the world. The dates when humans appeared in different places, and the paths they followed, are not entirely clear, but it seems plausible to assume that humans passed from North Africa through the Arabian and Sinai peninsulas to reach the Middle East about 60,000 years ago (ya), spread south and east to the Indian subcontinent and Asia about 50,000 ya, jumped from island to island of South Asia to reach Australia and neighboring land masses about 40,000 ya, crossed the Bering straight about 15,000 ya, and traversed the length and breadth of the Americas to reach Patagonia about 10,000 ya and Greenland about 4,000 ya. These migration paths and dates are shown in the figure below.
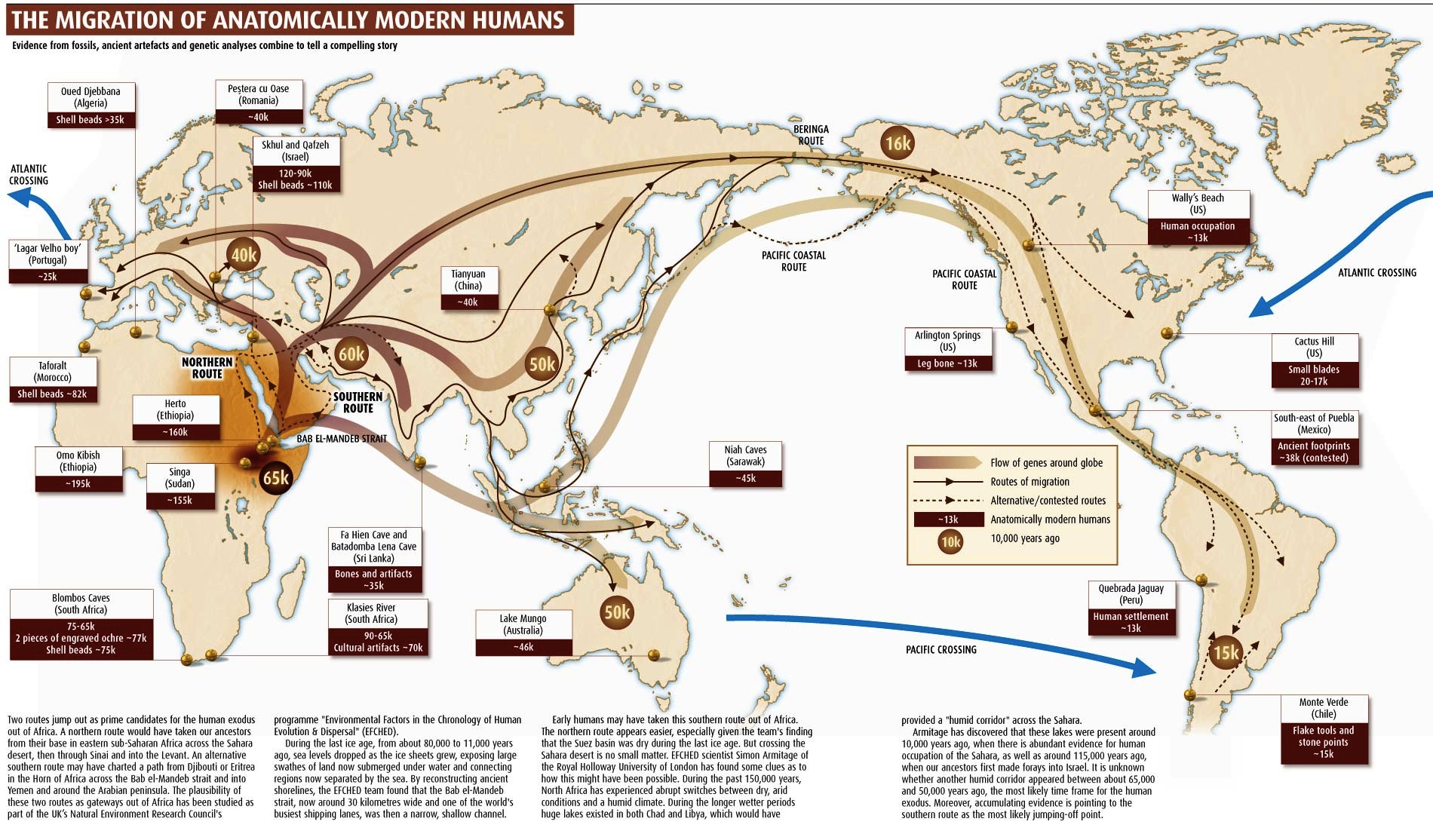
Figure 10. The migration of anatomically modern humans. Jones D, 2007. Going global: How humans conquered the world, New Scientist, October, 2007.
Recall that genetic distance is a measurement of how far the genomes of two organisms have diverged because of gene flow. Low genetic distance indicates relatively more gene flow, while high genetic distance indicates less gene flow. Thus, populations that freely mix are expected to have lower genetic distance and less genetic variation, while populations that are in some way isolated are expected to have higher genetic distance and more genetic variation.
Where and when they were able, humans constantly traveled in search of food or resources, for the purposes of trade, exploration or conquest, or because of disease or environmental pressure. This constant movement resulted in near constant gene flow, linking peoples from different parts of the world that were accessible to each other, and reducing the genetic variation between them. Hence, people from Europe and Central Asia appear to have mixed quite freely, despite the considerable distances involved, because the terrain they had to traverse did not restrict movement, producing relatively less genetic variation. In contrast, people from South Africa and Australia, or Papua New Guinea and the Amazon basin, must have mixed to some extent (the lengths of time involved make this inevitable), but remained sufficiently isolated by natural geographical barriers, such as the Sahara desert, the Asian archipelago and the Indian and Pacific oceans, that they remained somewhat genetically isolated and, over time, accumulated relatively more genetic variation. Differences in the relative ease of migration between different parts of the world, the extent of gene flow and genetic variation, are visible to us through the analysis of multiple genetic markers and sophisticated statistical techniques.
We can therefore answer the first of our broad questions about human genetic variation: what are the observable patterns of human variation? These patterns are as follows: the human genome shows gradual or clinal variation in areas where human have been able to move freely, with genetic distance proportional to the distances between them, and greater variation with discontinuous breaks between areas where significant natural geographical barriers limited gene flow for long periods of time. The American biologist, Noah Rosenberg, summarized this view in a study entitled, Clines, clusters, and the effect of study design on the inference of human population structure:
“If enough markers are used with a sufficiently large worldwide sample, individuals can be partitioned into genetic clusters that match major geographic subdivisions of the globe, with some individuals from intermediate geographic locations having mixed membership in the clusters that correspond to neighboring regions. For population pairs from the same cluster, as geographic distance increases, genetic distance increases in a linear manner, consistent with a clinal population structure. However, for pairs from different clusters, genetic distance is generally larger than that between intra-cluster pairs that have the same geographic distance. For example, genetic distances for population pairs with one population in Eurasia and the other in East Asia are greater than those for pairs at equivalent geographic distance within Eurasia or within East Asia. Loosely speaking, it is these small discontinuous jumps in genetic distance—across oceans, the Himalayas, and the Sahara—that provide the basis for the ability to identify clusters that correspond to geographic regions.”
If you would like to call these clusters races, then I would say you are free to do so. However, if you place several clusters together out of convenience, for example, “Africans,” or “Europeans,” or Asians,” or “Indians,” the categories become far less informative. For racial categories to have any meaning, at least in terms of genetic variation, you would have to say, “Bantu,” or “Lapp,” or Pathan” or “Bedouin,” or “Ainu,” or Mon Khmer,” or “Mayan,” bearing in mind, of course, that these groups themselves can undoubtedly be resolved with even greater precision.
I would like to draw attention to the first part of Rosenberg’s comments: “If enough markers are used with a sufficiently large worldwide sample.” The fact that we need to examine so many genetic markers, from so many people, with such powerful statistics, confirms what I stated above, namely, that differences between us are extremely small, and exist against a backdrop of overwhelming similarity. Nonetheless, broad patterns of genetic variation are clear. We can now turn to the second question posed above, “How were these patterns created?”
Forces that create genetic difference
Genetic difference is created by multiple forces working together over long periods of time. One force is the gradual accumulation of random mutations in populations with varying degrees of genetic isolation. Other forces involves processes similar to genetic bottlenecks. As I explained above, genetic bottlenecks, such as the Toba catastrophe, created our genetic similarity. These events can also create genetic difference. To understand how this is possible, consider a large herd of animals, such as gazelles. If a small group of gazelles leaves the main group and migrates to a new area, for example, by crossing to the opposite side of a mountain range, or penetrating a new valley, its genetic diversity, following the principles of a genetic bottleneck, will most likely be lower than that of the larger group. Likewise, the frequency of gene variants in the smaller group will most likely be different from that in the larger group. That is, not only will the smaller group contain less diverse genes, it will also contain a different assortment of genes. In this way, the smaller group will become different from the larger group. This process, by which a small group of individuals is separated from a larger group, reducing genetic diversity and increasing the genetic difference between between the smaller and larger groups, is known as the founder effect, commonly observed among species that colonize uninhabited territory, such as birds blown to distant islands, or fish suddenly separated in streams, lakes or drainage basins. The founder effect is perhaps best understood as a kind of genetic bottleneck which occurs when smaller groups are divided from larger groups.
Another process that can create genetic difference is known as genetic drift. Genetic drift describes the way that, in very small populations, random mutations or assortments of genes can have a significant impact on overall genetic composition. Once again, consider a large herd of gazelles. Within this herd there is a great deal of genetic diversity. If an individual is born with a particular assortment of genes, these genes will eventually be mixed, diluted, and subsumed by other genes in the herd. Over time, the gene frequencies in the herd will remain largely stable, unless they are effected by additional evolutionary forces, such as those described below. In fact, the gene frequencies in large groups can be mathematically quantified with some precision, following the principles of inheritance and Mendelian genetics. The important point is that, if a group is large enough, gene frequencies will remain relatively stable. Now imagine that a very small group is separated from the larger group. Naturally, individuals in the smaller group will have a particular assortment of genes. However, over time, these genes will not be mixed with other genes. Instead, the random assortment of genes present in the smaller group will predominate–their frequency will increase relative to the original population. In this way the two groups will become different. The effects of genetic drift are proportional to population size; in small groups, genetic drift can create substantial differences.
It’s easy to see how these processes can be extended to humans. As humans dispersed around the world, they traversed mountain ranges and deserts, crossed rivers and oceans, spread across plains and valleys. Each expansion of territory represented an opportunity for the founder effect to alter genetic composition. The gradual human settlement of the globe can be seen as a series of founder events, later tempered by varying degrees of gene flow.
This hypothesis is supported by evidence. Studies of genetic distance, such as those described above, consistently find that the genetic diversity of native people around the world is lower than the genetic diversity of native people in Africa. The entire global population, although it has grown enormously in size, is derived from a small founding population, or, rather, from many small founding populations. It follows genetic diversity around the world is lower than genetic diversity in Africa. This observation has several interesting implications that I will return to later.
Likewise, groups that remained relatively isolated over time, such as native people from Pacific Islands, or those from high mountain regions in the great ranges, such as the Himalaya or the Andes, or those protected by the Sahara desert or the Amazon rain forest, likely display some genetic differences due to genetic drift. These processes surely produced at least some of the genetic differences we observe today.
Before I go any further, I would like to emphasize that these processes are important insofar as they help us understand our history, and partially explain the small human genetic diversity that exists. However, they do not change the fact that humans are overwhelmingly more similar than they are different. Furthermore, they are not the only processes that create genetic difference. Instead, random genetic mutations, the founder effect and genetic drift provided the background diversity for other evolutionary forces to work. To understand these forces, we have to review the processes that drive evolutionary change.
There are two principal forces that drive Darwinian evolution: natural selection and sexual selection. I will describe each briefly, and discuss their relative contribution to human genetic variation.
Natural selection guides the development of traits that help living things survive in particular environment, such as, in the case of large mammals, powerful muscles and strong bones for catching prey or evading predators, thick fur to remain warm, refined senses and so on. The classic example, if only because it is so visually striking, is the elongated neck of the giraffe, that allows these creatures to browse contentedly on the upper canopy of trees, occupying their own ecological niche, free from other competitors. I should add, however, that in addition to things like strong bones and long necks, natural selection works equally to guide the development of more subtle traits, including behavior, such as herding instincts, maternal care and so on. Any traits that help living things survive in their environment will be maintained or magnified over time, for the simple reason that individuals which posses the genes that produce these traits will, on average, live longer and produce more offspring, and thus pass on these genes to successive generations, while individuals which do not posses these genes will not live as long nor produce as many offspring, subsequently their genes will not persist to the same degree over time.
Natural selection, then, helps produce living things that are well suited to their environment, and the natural world is full of myriad beguiling examples of this process at work, with endless variations and subtle complexities. The important point is that natural selection is about survival; for natural selection to operate, there must be some environmental pressure to shape a particular characteristic.
Sexual selection can be thought of as a unique counterpart to natural selection. Whereas natural selection guides the development of traits essential for survival, sexual selection guides the development of traits necessary for reproduction, regardless of whether or not these traits assist, or even hinder, survival. Any trait that help living things reproduce will be accentuated, because individuals which posses the genes that produce these traits will reproduce more and pass on more of their genes to successive generations, while individuals which do not posses these genes will reproduce less and pass on less of their genes to their offspring. In this case, the classic example is the peacock train, a great assemblage of feathers in bright hues that can be opened in a dazzling array. In a world in which sexual selection did not exist, biologists would be hard pressed to explain the peacock train. After all, the train requires a good deal energy to produce, maintain, and display, and it makes it difficult for peacocks to maneuver on the ground and take off and fly, and so makes peacocks vulnerable to predators. Based only on the principles of natural selection, it would seem that peacock’s feathers would grow shorter over time, much as giraffe’s necks have grown longer. Why do peacocks have trains? The explanation is that trains help peacocks reproduce; brilliant tail feathers are much admired by peahens, perhaps because they advertise the overall good quality of the peacock’s genes, as if to say, “I have managed to survive despite my beautiful train.” Thus, over time, peacocks with long train producing genes reproduced more, and passed on more of these genes to successive generations, while peacocks without long train producing genes, or peacocks with slightly shorter train producing genes, reproduced less, and passed on less of their genes to their offspring.
That is how sexual selection operates. I should say, again, that the traits on which sexual selection might exert a directive force are not limited to physical characteristics such as tail feathers; sexual selection can work equally on behavioral traits. Studying the elaborate, sometimes comical, often mystifying mating habits of various species is one of the most intriguing and entertaining aspects of biology.
What role, then, do natural and sexual selection play in creating patterns of human variation? First, I think it’s necessary to say that many, if not most, characteristics of humans, like the characteristics of all living things, must be the result of the combined effects of natural and sexual selection. For example, to single out some of the traits that make us distinctly human, our upright stance, large brains, complex language, and appreciation of music and art, must be the result of these evolutionary processes. Asserting that human characteristics are largely a product of natural and sexual selection, however, is not the same as asserting that patterns of human variation are produced in the same way. In the first case, it’s easy to appreciate that large brains conferred a survival advantage to our ancestors. In the second case, one would have to prove 1) that brain size varies in a systematic way in different parts of the world, 2) that the environment varies in the same way, and 3) that the environment exerted a differential selection pressure on brain size. This is a clearly more difficult.
Bearing this distinction in mind, are there any patterns of human variation that appear to be the product of natural selection? Yes. There are two: skin color and height. Skin color is the most obvious example. I have explained how skin color varies around the world (people from equatorial and tropical regions generally have darker skin, while people from temperate and polar regions have lighter skin). Here I will explain why skin color varies this way.
Skin color is related to the amount of melanin produced by specialized cells in the lowest layer of the epidermis. More melanin produces darker pigmentation, while less melanin produces lighter pigmentation. Melanin, in turn, absorbs ultraviolet radiation, and therefore protects humans from the damaging effects of the sun. Darker skin provides protection from harmful ultraviolet radiation, while lighter skin allows enough sunlight to penetrate to produce vitamin D. Skin color can thus be seen as a balance between protection from ultraviolet radiation and Vitamin D production. It follows that, if a native of the Indian subcontinent, for example, were transplanted to the north pole, they would most likely develop a vitamin D deficiency, while a Native Greenlander forced to live in the tropics would have a greater chance of developing skin cancer. This is what we observe. Australia, a continent in a tropical region, populated largely by immigrants from a temperate region, has one of the highest rates of skin cancer in the world. The variation of skin color with latitude is consistent, allowing for some variation, probably the result of migration, and for discontinuous breaks in landmasses.
In the case of height, the relationship is equally simple: native people from jungle and forest regions tend to be shorter, presumably because their small stature allowed them to move through and survive in dense undergrowth, while people from savanna and plains regions tend to be taller, perhaps because greater height allowed for more efficient stride length, or greater heat dissipation, any number of other reasons. Regardless, the relationship is generally consistent, once again, allowing for some natural variation.
Here we arrive another central finding of modern science: apart from skin color and height, there are no regular patterns of human variation that are the product of natural selection. To make this point clear, I can redress several common misconceptions. Kinky hair, common to many people throughout the world, does not provide greater sun protection. The epicanthic fold, common to Asian people, does not allow greater visibility in bright light. To emphasize this point, I can invent more exaggerated examples, which would be humorous, if they were not matched by equally absurd racist beliefs. Black hair is not an adaptation to help to help Asian people hide at night, the hooked nose of Andean people is not an adaptation to help them smell snow in the mountains, and so on.
It might seem surprising that science has only been able to find two examples of patterns of human variation that are the product of natural selection, however, recall that the criteria are difficult to meet: one has to show that some characteristic varies throughout different parts of the world, that some aspect of the environment varies in the same way, and that the environment exerts a differential selection pressure on the characteristic in question.
To demonstrate how difficult this is to prove, we can consider fundamental characteristics, such as intelligence. If you asserted that people from one part of the word were demonstrably more intelligent than people from another part of the world, and you wanted to explain this observation, you would have to propose some difference in the environment that, over time, exerted a selection pressure on intelligence, and produced one group of more intelligent people. For example, suppose you claimed that native people from Siberia were more intelligent than native people from Amazonia.You might argue that that cold climate in Siberia forced people to rely on their intelligence to devise solutions to survive in cold temperatures, and that this drove the evolution of greater intelligence. You would also have to show that other equal or perhaps greater challenges posed by living in the Amazon did not exist. This is impossible. Every environment presents it’s own challenges to survival, and there is no plausible mechanism that would drive the evolution of greater intelligence in one area versus another.
The same is true of other fundamental characteristics, such as aggression, love, violence, ambition, criminality, sexual prowess, empathy, religious faith and so on. There are no plausible mechanisms by which natural selection could have produced regular patterns of variation in these traits, despite numerous attempts to articulate these mechanisms.
There is one more point I would like to make before I move on. Changes in fundamental characteristics, produced by natural selection, in living things such as humans, with long lifespans and few offspring, take a relatively long time to emerge, probably many tens of thousands of years. The time spans involved are, at any rate, much longer than the time span in which we know that such changes would have had to occur to produce recognizable differences between groups of people around the world, specifically, the last 65,000 years, from the time when humans left Africa in large numbers until the present. Thus, even if there was a plausible mechanism by which natural selection produced regular patterns of variation, there has not been sufficient time for these changes to occur.
What about sexual selection? Having eliminated natural selection as a mechanism to produce patterns of variation, we are left with sexual selection. Here again science provides a clear answer: patterns of human variation are thought to be largely the result of sexual selection, though of a slightly different kind than that described above. As humans spread around the world, there was some natural variation between people from different regions, for the reasons described above, such as random distribution, genetic bottlenecks and so on. Then, as now, people would generally find attractive and favor the traits of their own group, and therefore reproduce within that group, accentuating these traits, with, of course, considerable mixing and gene flow in areas where contact with other groups was possible. This produced a mosaic of different sexually selected physical characteristics, such as facial and body features, hair type and color, eye shape and color, even height and skin color, operating within the boundaries imposed by natural selection.
An important point is that, in contrast to fundamental characteristics, superficial physical characteristics, shaped by sexual selection, can change relatively quickly, certainly within the 65,000 year period that patterns of variation emerged. Therefore, we have clear mechanism through which patterns of variation in superficial physical characteristics emerged, and sufficient time for these changes to occur.
I mentioned that sexual selection in humans is somewhat different from sexual selection in other creatures. We are defined and created by culture, and culture exerts its own selective force. For example, mating preferences are strongly influenced by language, religion and local customs. These cultural preferences constitute a form of sexual selection, and play an important part in creating patterns of human variation. We return to Dawkins in The Ancestor’s Tale:
“The fact that we depart so strongly from random mating in directions determined by language, religion, and other cultural discriminators, has done very odd things to our genetics in the past. Even though, if you take the totality of genes into account, we are a very uniform species, we are astonishingly variable in superficial features which are trivial but conspicuous.”
I showed why it is difficult or impossible to propose a plausible mechanism by which natural selection could have produced patterns of variation in fundamental traits. I will do the same for sexual selection, and thereby make another point about evolutionary processes. Consider aggression. It’s entirely reasonable to imagine that in one group a trait such as aggression was favored by sexual selection. Individuals who were more aggressive were able to mate more often, while individuals who were less aggressive were not able to mate as often. Over time this group would have grown more aggressive. However, if you proposed this mechanism to explain why one group was more aggressive than another, you would have to explain why aggression was not equally favored in other groups. This would be very difficult, because various forms of competitive rituals, such as sports, and violent behavior, such as murder and war, are universal.
The same is true for the other fundamental characteristics; there are no good reasons to suppose that one group or culture would have developed these traits while others did not. Thus, while our superficial physical traits changed over time, our fundamental characteristics did not.
Individual variation and environmental effects
It should be clear that my arguments are directed as refuting the notion that there are regular differences in fundamental characteristics between groups of people around the world. However, even if such differences did exist, there are powerful reasons why they would be 1) nearly impossible to find, and 2) essentially meaningless. The explanation for why this is true is related to two important ideas: individual variation and environmental effects.
So far, I have used examples from the animal world, such as large mammals, to illustrate my points. Here, I will use plants, simply because the figures are so easy to understand. I will then extend the same arguments to human variation.
The first figure below shows two groups of plants, a shorter group and a taller group. As you can see, the average height of the shorter group (9.90 cm) is slightly lower than the average height of the taller group (10.35 cm). However, the difference between the average heights of the two groups is much smaller than the differences between the heights of individual plants within each group. This is true of nearly any characteristic we might choose to study in plants or animals; individual variation is almost always much larger than average differences between closely related populations living in similar conditions.
The second figure below also shows two groups of plants, one grown in nutrient poor conditions, and the other grown in nutrient rich conditions. The average height of the nutrient poor group (3.75 cm) is much lower than the average height of the nutrient rich group (20.95 cm). In this case, differences in height created by environmental conditions are much greater differences caused by individual variation within one or another group. It would be impossible to discern differences between groups, because you would not be able to tell whether they were created by underlying genetic variation or the presence of absence of nutrients. Again, this is true of many characteristics in plants or animals; the effects of the environment are powerful, and hide much weaker individual and group variation.
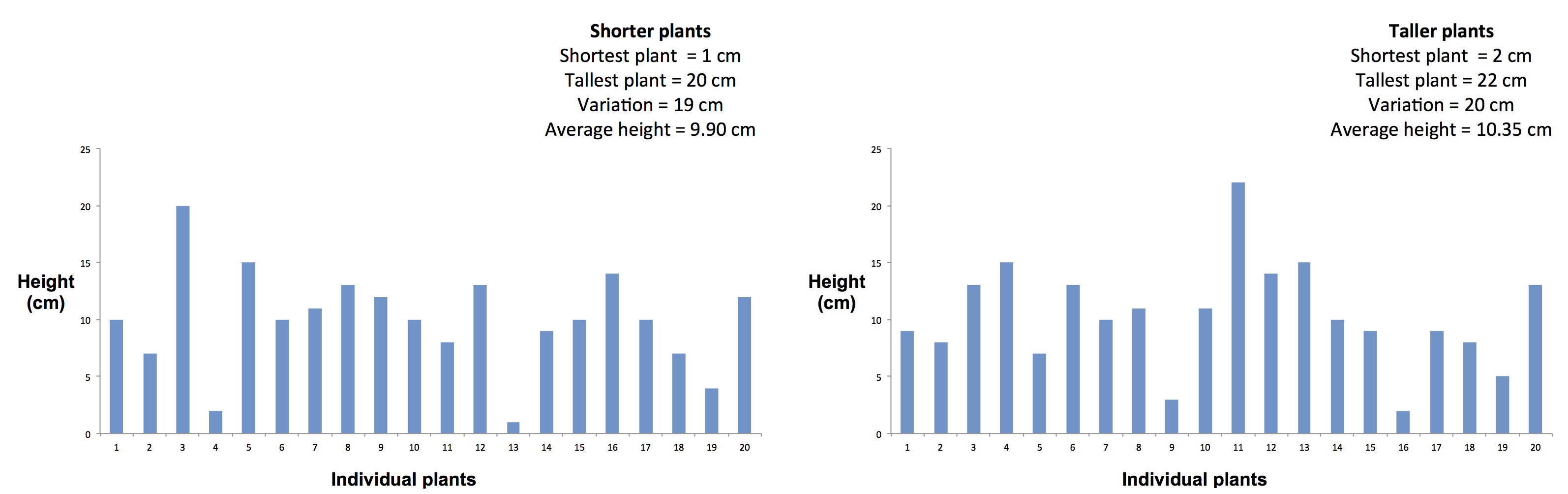
Figure 11. The effect of individual variation on plant height.

Figure 12. The effect of environmental conditions on plant height.
How can these ideas be extended to human variation? For the sake of argument, and contrary to what I explained above, let’s assume there are differences in fundamental characteristics between people around the world. I already presented examples relating to intelligence and aggression, so let’s consider mathematical ability, although we could equally discuss linguistic ability, fluency with abstract symbols, or any number of other physical or mental characteristics. Imagine that you devised some sort of test for mathematical ability, and found a small difference between two groups, for example, a difference of one percent or a fraction of one percent. As above, this difference would be much smaller than the individual differences in mathematical ability between members of the same group. Recall your own mathematics classes; surely, the grades of all the students in your classes did not vary by one percent but five, ten, twenty percent or more.
In similar fashion, imagine that you applied your mathematics test to another two groups, one in an area with little or no educational opportunities, and the other in an area with a well-developed school system. Obviously, people with little or no exposure to math, or education of any kind, would perform poorly on a conventional test of mathematics, compared to people who spent many years in school.
Hopefully my point is clear; even if differences in fundamental characteristics between people around the world did exist, they would be absolutely overwhelmed by individual variation within any group, and by differences in environmental conditions between groups. This is why I don’t believe we will ever find such differences, and why, if we do, they will be irrelevant to any meaningful discussion of human behavior or achievement.
Conclusions
We have traversed a great number of interrelated subjects, so I think it’s appropriate to summarize our main conclusions:
- All humans belong to a single species, Homo sapiens. We know this because all humans can and do breed successfully.
- All humans are very similar genetically, far more similar than many other animals, including most large mammals and primates.
- The average human global FST varies between about 0.05 and 0.15. This is much smaller than accepted thresholds to divide species into sub species, the biological term synonymous with race. Therefore, there is no scientific basis to distinguish human races.
- The similarity between humans was most likely caused by a genetic bottleneck that occurred about 70,000 years ago, during the Toba catastrophe, before humans began to disperse around Earth in large numbers.
- Despite our genetic similarity, there are regular patterns of variation between native people around the world. We know this because inter-observer correlation regarding such variation is high.
- If we study large numbers of genetic markers from large numbers of people, we find that humans can be divided into groups associated with geographical areas separated by great distances and formidable natural barriers, such as continents, oceans, mountains, and deserts. These barriers created varying degrees of genetic isolation. Humans always migrated, and gene flow always occurred, but some areas remained sufficiently isolated that differences developed.
- The differences between groups of native people developed in the last 65,000 years, from the time when humans began to leave Africa in large numbers until the present. This is a very short time in terms of evolutionary change.
- The differences between groups of native people were produced by several processes, including natural selection, acting globally, over long periods of time, and sexual selection, acting locally, over short periods of time.
- The differences between groups of native people are limited to superficial physical characteristics, such as skin color, height, and facial and body characteristics. There is no evidence that there differences in fundamental characteristics. Furthermore, there are good reasons why we don’t think we will ever find these differences.
- The reasons we don’t think we will ever find these differences are 1) these differences exist against a backdrop of overwhelming genetic similarity, 2) there has not been enough time for these differences to develop, 3) there is no plausible selection mechanism that could have produced these differences, and 4) individual variation and the effects of environmental conditions are much greater than average differences between groups.
These conclusions show that the racist views held by so many people, including scientists, for hundreds of years, are demonstrably false. There are no human races. They also show that the views held by scientists in the recent past are incomplete. There are recognizable differences between native people around the world, although these differences are very small, and limited to superficial physical characteristics.
The findings of modern science have great explanatory power, and are supported extensive evidence from archeology, anthropology, biology and genetics. In addition, they offer a wide variety of hypotheses that can be tested, and either verified or abandoned, as new evidence becomes available. I hope that I have explained this information as clearly as possible, and established a strong scientific basis for racial equality.
